Padraig O’Kiely, Aidan Moloney, Thomas Keating and Patrick Shiels
Teagasc, Grange Teagasc (Irish Agriculture & Food Development Authority) Centre, Dunsany, Co. Meath
END OF PROJECT REPORT
MAY 1999
ARMIS No. 4283
ISBN 1 84170 046 0
Teagasc acknowledges with gratitude the support of European Union Structural Funds (EAGGF) in financing this research project
European Agricultural Guidance and Guarantee Fund
Beef Production Series No. 10
This report summarises a series of experiments, which are grouped under the following headings:
1. REPLACING OLD PERMANENT GRASSLAND WITH RYEGRASS SWARDS - IMPACT ON SILAGE-BASED BEEF PRODUCTION
A comprehensive research programme showed the potential benefits of replacing a productive old permanent grassland pasture dominated by indigenous species with new perennial or Italian ryegrass swards, when each was managed intensively, conserved as silage and fed to beef cattle. Ancillary experiments showed how the silage systems could be modified to improve productivity. However, they also showed that under a less intensive regime, replacing this old permanent pasture by ryegrass reseeds would be difficult to justify.
Importance of grassland
The Irish landscape has been alternately dominated by grassland and deciduous forest over the past 13000 years, and our current large expanses of grassland (about 90% of farmland) have largely developed over the past three centuries. We are unique within the European Union in that over 90% of our ruminant feed is derived from grazed or conserved permanent grass.
A wide range of vegetation types occur in our grassland, with botanical composition in any location depending on environmental and management factors. Permanent grassland dominated by indigenous (unsown) species is common, and can be an important ecological resource for diverse flora and fauna. Such grassland also exhibits a considerable range in commercial agricultural characteristics such as annual quantity and pattern of herbage production, nutritive value, ensilability and trafficability.
A small but cumulative replacement of old permanent grassland by permanent swards of perennial ryegrass (Lolium perenne) has taken place on many farms in recent decades, such that currently about half the silage in the country is conserved from ryegrass-dominant swards. This replacement has often occurred as part of farm management strategies to improve incomes by increasing the output of saleable meat or milk per hectare of grassland. The commercial benefits accruing from replacing old permanent grassland swards by perennial ryegrass swards depend largely on the productivity of the former and the ability of the subsequent management practices to maintain the new ryegrass swards and derive full benefit from them.
Research programme
A research programme was undertaken to develop optimal management strategies for three sward types used for the intensive production of silage for beef cattle. The swards were an old permanent grassland sward that had been intensively managed for several decades and had not been reseeded in living memory (
Table 1.1), a mid-season diploid perennial ryegrass and finally a tetraploid Italian ryegrass (Lolium multiflorum) as a specialist silage crop. Specifically, the aims of the four components of the research programme were to:Experiment 1.1. Beef carcass output
Method. Replicate main plots within 9 ha of old permanent grassland were left unchanged or reseeded in early September with perennial or Italian ryegrass. Average dates for the four harvests of the old permanent grassland and perennial ryegrass swards in each of the following three seasons were May 25, July 9, August 28 and October 27, while the average for the five harvests per year of the Italian ryegrass sward were May 10, June 15, July 19, August 27 and October 26. Each sward received 430 kg fertiliser N/ha each year, and no animal manures were spread. Grass was harvested without wilting and was weighed into silos and all silage weighed from silos, so as to measure conservation efficiency. In year 1, finishing steers were offered ad libitum the four or five harvests of silage combined, in sequence, for 126 days, supplemented with 2.0 kg concentrates per head daily. In year 2, finishing heifers were offered ad libitum the silages from the first growth, second growth or remaining growths combined of the three swards for 103 days. The grass harvested in year 3 was not fed to cattle as part of this experiment.
Results. Although the average yield benefits in favour of replacing this old permanent grassland sward by ryegrass swards were small (especially with perennial ryegrass), there were benefits in terms of both digestibility and ensilability (
Table 1.2). There was also a considerable effect of the prevailing weather each year on the yield, digestibility and ensilability of each sward. The differences in ensilability indices were reflected in average pH values of all silages made from old permanent grassland, perennial ryegrass and Italian ryegrass swards of 4.19, 4.00 and 3.82, respectively. Correspondingly, forage digestibility decreased by 52, 20 and 47 g/kg during ensilage, and the mean recovery rates of edible silage DM were 781, 781 and 750 g/kg (2 years only). Although neither silage DM intake nor carcass gain was affected by sward type in year 1, in year 2 the Italian ryegrass supported higher intakes and carcass gains with both first and second cuts, as did perennial ryegrass for the first cut, compared to the old permanent grassland sward.When the yield, digestibility, conservation, silage intake and animal performance data were combined, the results (
Table 1.3) showed a benefit to reseeding in terms of output of beef carcass per ha.Experiment 1.2. Conservation losses
Method. In the third season after reseeding, precision-chop grass at each harvest date was ensiled in replicated 2 m high laboratory silos. No silage additive was used.
Results. All silages underwent lactic acid dominant fermentations, but the efficiency and ease of fermentation was better for the ryegrasses (
Table 1.4). However, poorer recoveries of silage DM occurred at silo opening for the Italian ryegrass crop, due in part to greater losses of effluent. Subsequent to silo opening, silages made from the old permanent grassland sward were slower to heat and deteriorate than those from the ryegrass reseeds.Experiment 1.3. Nitrogen fertiliser
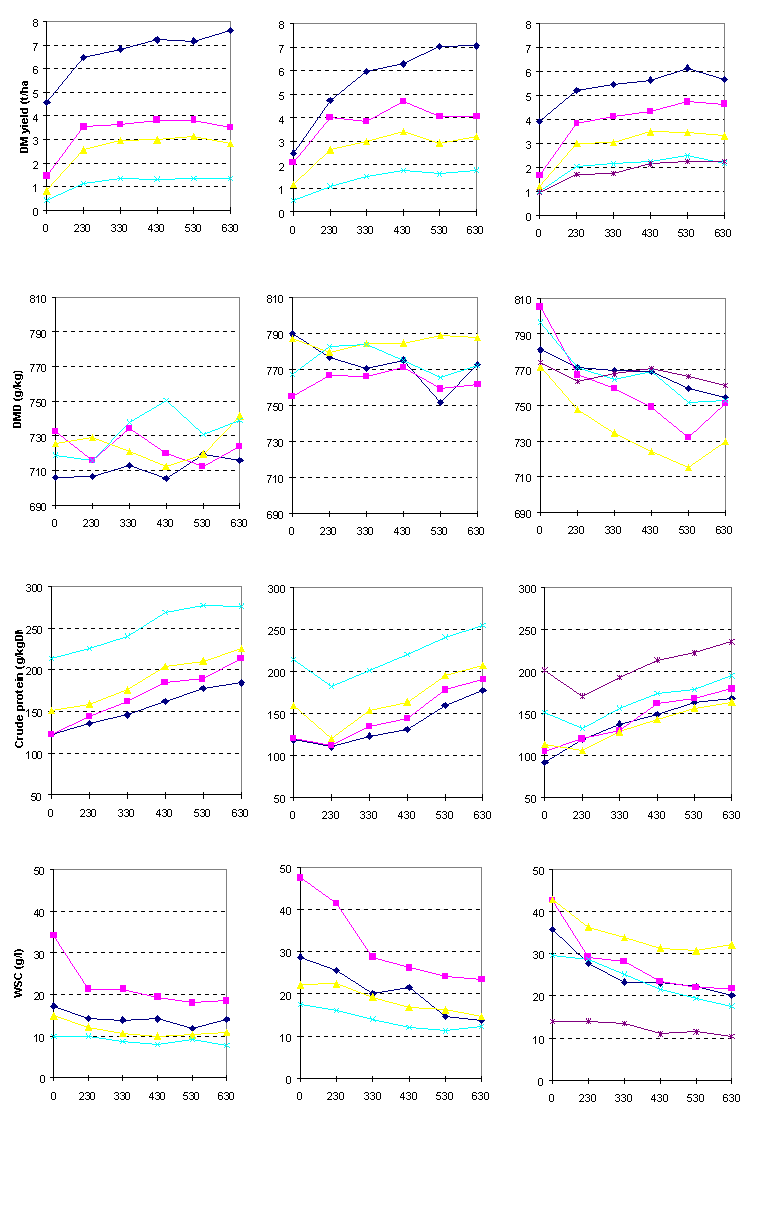
Method. After the first year of the beef carcass output experiment, plots located within the old permanent grassland pasture were either left unchanged or were reseeded with perennial or Italian ryegrass. In the first and second years after this reseeding, sub-plots were treated with 0, 230, 330, 430, 530 or 630 kg N per hectare each year. These were harvested on the same dates as the large field (i.e. main) plots.
Results. Neither sward botanical composition nor tiller density were altered by the rate of fertiliser N applied. In the absence of fertiliser N being applied, mean annual DM yields were 7.3, 6.2 and 8.7 t/ha, mean in vitro DMD values were 721, 775 and 786 g/kg, mean crude protein concentrations were 153, 153 and 132 g/kg DM and mean WSC concentrations were 19, 29 and 33 g/kg aqueous phase. Increasing rates of fertiliser N application generally increased DM yield and crude protein concentration, and decreased WSC concentration (Figure 1.1) - the WSC depression was most evident with ryegrasses.
Increasing rates of N lead to a fairly consistent decrease in the DMD of Italian ryegrass, but not of perennial ryegrass or the old permanent grassland sward.
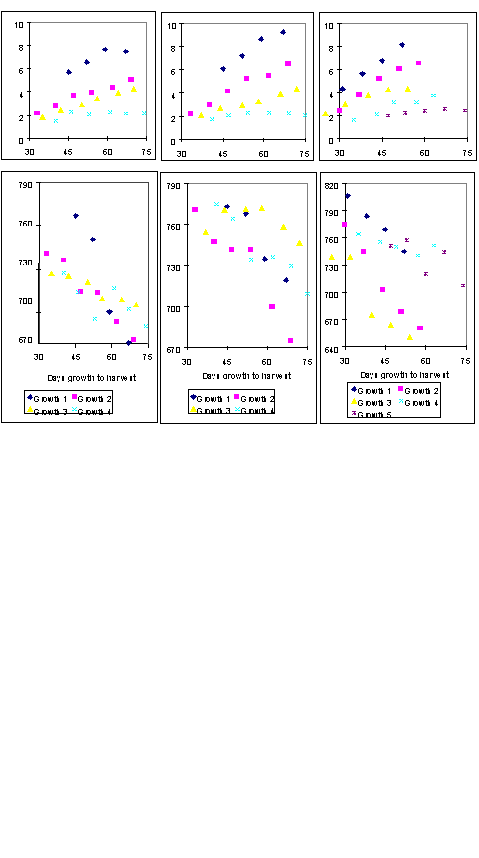
Experiment 1.4. Harvest date
Method. In the second and third years after reseeding, plots located within each growth of each sward had sub-plots harvested at weekly intervals before and after the harvest dates of the main plots.
Results. There was a major effect of year on the results. Averaged across all 3 swards and all harvest dates, the mean DM yields in years 2 and 3 were 19.25 and 15.97 t/ha, respectively, with corresponding values for DMD of 725 and 738 g/kg, crude protein of 176 and 215 g/kg DM and WSC of 96 and 80 g/kg DM. Delaying harvest date generally increased DM yield and decreased both DMD (Figure 1.2) and crude protein concentration. The effects on WSC concentration were less clearcut due to the over-riding effects of prevailing weather.
Conclusions
2. BACTERIAL INOCULANTS UNDER DIFFICULT ENSILING CONDITIONS
Lack of control of fermentation; The fermentation process that preserves grass as silage is controlled by a complex interacting matrix of factors. These include
Because most of these factors are not fully under the farmers control, the silage fermentation process is relatively uncontrolled compared to industrial fermentations involved in the manufacture of enzymes, alcohol, yoghurt etc. Farmers try to follow a package of practices that predispose silage fermentation to dominance by lactic acid bacteria, but the precise extent and intensity of this dominance cannot be assured in advance.
Silage-making conditions; Weather has an enormous direct and indirect impact on the ensilability of grass. This begins weeks or months before harvesting since it effects the timing of spreading inorganic or organic nitrogen and its rate and efficiency of conversion to crop dry matter (DM). In the days preceding and during harvesting the interaction of rainfall, sunshine and both night and day-time temperatures directly influence crop dry matter and sugar concentration, buffering capacity and microflora. At the same time, weather can have indirect effects by influencing harvesting speed and the risk of contamination of grass by soil. Thus, the ease of preserving grass as silage can vary widely due to factors substantially outside of the farmers control. It is therefore not surprising to see considerable variability in grass ensilability from week to week, season to season and year to year. Crops can range from those which will readily undergo an extensive lactic acid dominant fermentation through to those where dominance by lactic acid bacteria over clostridia or enterobacteria will be quite difficult to achieve.
Assuming that good silage-making practices prevail (i.e. quick and full achievement of anaerobic conditions and absence of contamination of forage by soil or animal manure), grasses with a high concentration of fermentable carbohydrate and a low buffering capacity are likely to be relatively easy to preserve as silage compared to grasses of low fermentable carbohydrate concentration and high buffering capacity.
Inoculants; Among the strategies taken by farmers to promote the dominance of silage fermentation by lactic acid bacteria are
The main types of additives have been those supplying inorganic or organic acid, sugar or lactic acid bacteria. Until recently, the main component of bacterial inoculants were homofermentative lactic acid bacteria, either alone or with the inclusion of fibrolytic enzymes. These have normally been applied to grass at ensiling at rates calculated to add between 100,000 and 1 million viable bacterial cells per gram of forage.
LITERATURE - GOOD SILAGE-MAKING CONDITIONS
Most of the published literature relating to the efficacy of bacterial inoculants as silage additives report experiments conducted under relatively favourable silage-making conditions, and where silages made without additives therefore underwent a lactic acid dominant fermentation. In these experiments, the addition of high numbers of viable homofermentative lactic acid bacteria frequently increased the rate of lactic acid accumulation in silage and thus the rate of decline in pH. The effects on final pH or ammonia-N concentration were not consistent, but often resulted in a small decrease in pH and ammonia-N concentration. When these silages have been fed to beef cattle or dairy cows, increased silage intake and improved animal performance have tended to follow where improvements in silage fermentation occurred.
DIFFICULT ENSILAGE CONDITIONS
Climatic conditions in Ireland frequently result in grass crops being difficult to preserve. A critical issue for Irish farmers is therefore whether or not the benefits accruing from the use of homofermentative lactic acid bacteria, frequently reported in the literature as prevailing under good silage-making conditions, also occur under more challenging ensiling conditions. Evidence on the efficacy of such inoculants as silage additives under difficult conditions is scarce, and is needed because of the frequency with which such conditions prevail in Ireland.
A series of experiments were conducted under difficult ensiling conditions to
(a) Silage conservation (Experiments 2.1 to 2.88)
A series of 88 experiments were conducted under difficult ensiling conditions at Grange Research Centre to investigate the efficacy of a range of silage inoculant additives based on supplying relatively high numbers of homofermentative lactic acid bacteria. In each experiment, unwilted grass from Lolium perenne dominant permanent grassland swards was ensiled in laboratory silos (O’Kiely and Wilson, 1991; 4 silos per treatment) either
Grass crops generally received cattle slurry plus 100 to 125 kg N/ha, and were shaded by a plastic canopy for 48 h prior to harvesting. In addition, silos were not sealed until the day after harvesting and additive treatment, to simulate the rate of filling and sealing of farm silos.
The mean (sd) composition of the herbage at ensiling was 156 (25.1) g DM/kg, in vitro DM digestibility (DMD) 733 (40.0) g/kg, 213 (41.2) g crude protein/kg DM, 134 (29.0) g ash/kg DM, 14 (6.1) g WSC/l aqueous extract and buffering capacity 352 (85.2) mEq/kg DM.
In general, silages made without additive in these experiments had considerable clostridial activity (
Table 2.1).Conclusion: Whereas conservation characteristics were improved by the rate of formic acid applied, the inoculant treatment did not result in significant improvements (neither in individual experiments or when averaged across 88 experiments).
(b) Interaction of additive with concentrates when offered to beef cattle (Experiments 2.89 - 2.90)
Two experiments were conducted to determine the effects of a homofermentative lactic acid bacterial inoculant or formic acid, applied to grass under difficult ensiling conditions (
Table 2.2), on the nutritive value of silage offered to beef cattle. Furthermore, the interaction of these effects with the feeding of supplementary concentrates was investigated.These experiments were carried out under very difficult ensiling conditions and therefore yield some very important results for Irish silage-makers (Tables
2.3, 2.4, 2.5 and 2.6). A considerable amount of digestion and animal metabolism supporting data were also produced in these experiments.Conclusions from all these data are:
Experiment 2.89
Experiment 2.90
(c) Conservation and nutritive value of silages made using lactic acid bacteria selected from Irish silages
Two experiments were conducted, the objectives of which were:
Grass that was difficult to preserve (
Table 2.7) was harvested unwilted using a precision-chop forage harvester. Alternate pairs of loads of grass were allocated to the following treatments in the following order:(i) Untreated control
(ii) Lactobacillus plantarum (Ecosyl, Zeneca Bioproducts and Fine Chemicals plc.) applied at 3.06 l/tonne
(iii) Lactobacillus plantarum (DCU 101) applied at 3.06 l/tonne
(iv) Pediococcus spp. (G24) applied at 3.08 l/tonne
(v) Lactobacillus plantarum (DCU 101) + Pediococcus spp. (G24) applied at 3.01 l/tonne
Data presented for these experiments in Tables
2.8, 2.9, 2.10, 2.11, 2.12 and 2.13 involve the first farm-scale results recorded for two new strains of bacteria used as silage inoculants. Ensiling conditions were relatively difficult as predicted from crop conservation data.Conclusions which can be drawn are
Experiment 2.91
Experiment 2.92
(d) Lactic acid bacteria + selective sterilant (Experiments 2.93-2.95)
Published data by Kalzendorf et al. (1991) indicated the potential of sodium formate as a selective sterilant against Enterobacteria and Clostridia but to which lactic acid bacteria were relatively tolerant. The objectives of these experiments were:
Experiment 2.93. Inclusion of a selective sterilant with lactic acid bacteria added to forage at ensiling
Introduction. Domination of silage fermentation by lactic acid bacteria depends on the interaction between a range of factors, including the supply of available fermentable substrate, buffering capacity and the numbers, types and activity of desirable lactic acid bacteria and other undesirable micro-organisms. Addition of homofermentative lactic acid bacteria to forage at ensiling sometimes stimulates lactic acid production, while sterilants have been added to depress or inhibit microbial activity, particularly that of undesirable micro-organisms.
Kalzendorf et al. (1991) stored lactic acid bacteria (LAB; lactobacilli and streptococci) in a range of potential sterilants and found the best revival ability rate for LAB was when stored in sodium formate, even at concentrated solutions. The objective of this study was therefore to determine the effects of adding L. plantarum and sodium formate, either separately or together, on the pattern of silage fermentation.
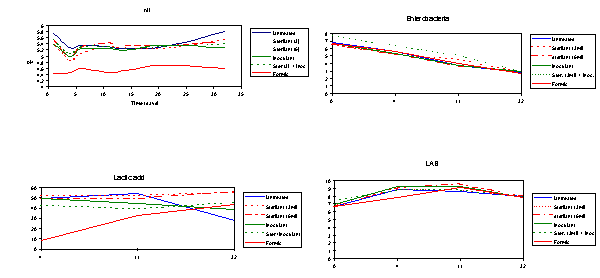
Materials and Methods. A Trifolium repens dominant sward, to which liquid cattle slurry had previously been applied, was chopped through a precision chop forage harvester and ensiled in laboratory silos (6 kg/silo) following treatment with (A) no additive, (B) formic acid (850 g/kg; 3 ml/kg forage), (C) sodium formate (saturated solution; 3 ml/kg), (D) sodium formate (6 ml/kg), (E) L. plantarum (Ecosyl, Zeneca Bioproducts and Fine Chemicals Ltd., 106 CFU/g fresh crop) or (F) L. plantarum plus sodium formate (3 ml/kg). Twelve silos were prepared per treatment and four were opened on days 4, 11 and 32 post ensiling. pH values were determined in effluent collected from all silos on days 1, 4, 6, 11, 14, 21 and 32 post ensiling. At silo opening, microbiological and chemical composition of the silages were determined.
Results. Figures 2.1 to 2.4 summarise some of the chemical and microbiological dynamics. Harvested forage had mean (s.d.) dry matter (DM), water soluble carbohydrate (WSC), in vitro DM digestibility (DMD), buffering capacity and nitrate concentrations of 110 (1.0) g/kg, 20 (5.8) g/kg aqueous extract, 660 (8.6) g/kg, 751 (18.4) m.equiv/kg DM and 890 (47.3) ppm aqueous extract, respectively. Microbiological analysis of the crop at harvest revealed high numbers of Enterobacteria (5.12 x 106 CFU/g fresh crop) and Clostridia (3.6 x 104 CFU/g fresh crop), but low numbers of lactic acid bacteria (3.53 x 104 CFU/g fresh crop). In a separate dose response study, sodium formate was shown to have no inhibitory effect on lactic acid bacteria but to inhibit Enterobacteria. The mean chemical and microbiological composition of the silages after 32 days ensilage is shown in Table 2.14.
Conclusions.
Although all silages were of poor fermentation quality after 32 days ensilage, formic acid did give some improvement, and a higher application rate was necessary. Sodium formate (3 and 6 ml/kg) and L. plantarum and L. plantarum plus sodium formate (treatments C to F) increased the rate of pH fall to day 4, at which stage WSC appeared to become limiting. Both rates of sodium formate selectively inhibited saccharolytic clostridial activity, as did L. plantarum and L. plantarum plus sodium formate to a lesser extent.Experiments 2.95 and 2.96. Combining sodium formate with a Lactobacillus plantarum inoculant applied to herbage at ensiling
Introduction: The ability of added lactic acid bacteria (LAB) to improve silage fermentation depends on the application rate and nature of the added product, the chemical, microbiological and physical characteristics of the crop being ensiled, and on the silage-making and storing conditions that prevail. Inoculation of difficult-to-ensile grass with homofermentative LAB at ensiling frequently fails to improve conservation characteristics (Shiels et al., 1993). Although Kalzendorf et al. (1991) and Weissbach et al. (1991) proposed that the inclusion of sodium formate with Lactobacillus plantarum applied to grass at ensiling could selectively inhibit the activity of undesirable micro-organisms such as Enterobacteria and Clostridia but not have a detrimental effect on the activity of LAB, Shiels et al. (1995) failed to obtain either an additive or synergistic effect of added L. plantarum and sodium formate on the conservation of difficult-to-ensile grass. The objectives of the present experiments were to determine the effects of adding L. plantarum and sodium formate, either separately or together, on the fermentation pattern of two forages of contrasting ensilability to that reported by Shiels et al. (1995), and to compare these effects to those of formic acid.
Materials and Methods: In Experiments 2.95 and 2.96 respectively, forages were harvested in early December from a Lolium perenne sward recently sprayed with soiled water and from a Trifolium repens dominant grassland sward in early May. On each occasion forage was harvested at a stubble height of 5 cm using a reciprocating mower and precision-chopped. In Experiments 2.95 and 2.96 respectively, sixteen and twelve samples of grass (each 7 kg) were randomly allocated to each of the following treatments: (A) no additive (negative control), (B) sodium formate, (C) L. plantarum (Ecosyl, Zeneca Products and Fine Chemicals Ltd.), (D) L. plantarum plus sodium formate, and (E) fomic acid (850 g/kg; Add-F, BP Chemicals Ltd.; positive control). Each additive treatment was applied manually immediately prior to ensiling, at 3 ml/kg forage. Sodium formate was applied as a saturated solution while L. plantarum was applied at a rate targetted to apply 106 colony forming units (CFU)/g forage. For treatment D, the lyophilized L. plantarum was reconstituted in the sodium formate solution. Forages samples were then stored overnight to permit some respiration occur, and 6 kg (excluding additive) sub-samples ensiled in laboratory silos. Silos were stored at ambient environmental conditions and at approximately 15oC. Four silos per treatment were opened on days 7, 28, 60 and 100 of ensilage in Experiment 2.95 and on days 7, 28 and 60 in Experiment 2.96.
Results: The mean (s.d.) composition of the harvested forage in Experiment 2.95 was dry matter (DM) 145 (9.5) g/kg, in vitro DM digestibility 712 (16.6) g/kg, crude protein 234 (1.7) g/kg DM, water soluble carbohydrates (WSC) 6 (1.2) g/L, buffering capacity 466 (7.2) mEq/kg DM and lactic acid bacteria (LAB) 3.65 x 104 CFU/g. The corresponding values in Experiment 2.96 were 172 (7.0) g/kg, 843 (9.9) g/kg, 182 (7.4) g/kg DM, 23 (2.8) g/L, 595 (22.0) mEq/kg DM and 3.30 x 103 CFU/g. LAB numbers enumerated after storing the remaining additive for treatment D for 24 h suggested that sodium formate had relatively little effect on the viability of the L. plantarum inoculant in either experiment. Silage composition data are summarised in Table 2.15.
Discussion: Ensilage conditions were not as severe as in the experiment reported by Shiels et al. (1995). Silage made without additive preserved better in Experiment 2.96 than Experiment 2.95. L. plantarum applied alone had no impact on fermentation under the low WSC supply conditions in Experiment 2.95, but promoted a more dominant homofermentative LAB fermentation under the higher WSC regime in Experiment 2.96. Sodium formate applied alone had little impact on fermentation and did not prevent silage preserving poorly. The inclusion of sodium formate did not enhance the effectiveness of added L. plantarum. Formic acid restricted fermentation and improved preservation.
Conclusion. Combining sodium formate with a L. plantarum inoculant applied to herbage at ensiling did not produce an effective preservative treatment. Formic acid was an effective preservative.
References. Kalzendorf, C., Kwella, M. and Weissbach, F. (1991). Landbauforschung Volkenrode, Sonderheft 123: 442-446. Shiels, P., Hayden, S., Doyle, E., O’Kiely, P. and Caffrey, P.J. (1995). Irish Journal of Agricultural and Food Research, 34: 83. Shiels, P., O’Kiely, P., Moloney, A.P. and Caffrey, P.J. (1993). Irish Journal of Agricultural and Food Research, 32: 95. Weissbach, F., Kalzendorf, C., Reuter, B. And Kwella, M. (1991). Landbauforschung Volkenrode, Sonderheft 123: 273-282.
Overall Conclusions to Section 2
The data presented in this part of the report are unique in that four farm-scale experiments were carried out, all of which examined the effects of homofermentative lactic acid bacterial inoculants on the fermentation and nutritive value of difficult-to-ensile herbage (i.e. wet crops with low WSC concentration and high buffering capacity). Herbages of the type ensiled in these experiments have been the grey area to date because inoculant effects on silage preservation have not previously been examined in any great detail. The suggestion that positive benefits could be achieved from the use of bacterial inoculants under such challenging conditions has caused much debate. On the basis of the data presented here, homofermentative lactic acid bacterial inoculants could not be recommended to effect a successful preservation in difficult-to-ensile crops. Such an observation agrees with Rooke et al. (1990) who proposed that if there is insufficient substrate available from which acid can be produced, then currently available LAB silage inoculants are unlikely to produce silage with a sufficiently low pH to prevent the proliferation of undesirable micro-organisms and subsequent spoilage.
Conclusions that can thus be drawn regarding the conservation and feeding value of difficult-to-ensile herbage are:
3. SULPHITES TO IMPROVE AEROBIC STABILITY
Sulphites have been used in the human food industry to inhibit aerobic deterioration initiated by yeast. Although silage is stored anaerobically in silos, the silage is subsequently exposed to air at feedout, both at the silage face in the silo and in the feed-manger.
The present series of experiments studied the effects of sulphites
(a) added to silage at feedout on aerobic stability - Experiments 3.1 to 3.6
(b) added to an in vitro rumen fermentation medium on the extent of digestion - Experiment 3.7
(c) added to forage at ensiling on subsequent aerobic stability at feedout - Experiments 3.8 to 3.12
Experiments 3.1 to 3.6 Aerobic stability of unwilted silages treated with additives at feed-out
Introduction. Quantitative and qualitative losses from silage due to aerobic microbial activity at feed-out may be considerable. Such activity is frequently initiated by lactate utilising yeast, which may eventually be succeeded by moulds. Sulphites and sodium chloride have been used to preserve foods destined for human consumption from comparable aerobic activity. The objective of the present experiments was to determine the effects of applying sulphites and sodium chloride at feedout on the aerobic stability of a range of unwilted silages.
Materials and Methods. Aerobic stability was assessed by placing 6 kg silage in polythene-lined polystyrene (2.5 cm thick) boxes (59 x 39 x 22 cm) with a polystyrene lid loosely fitted and stored at 20oC. Temperature and pH were monitored daily. Silage temperature was compared with those of containers of water held in the same environment (reference temperature). If either pH or temperature failed to rise during the assessment, an interval period one day longer than the assessment was assumed to be the interval preceding a pH or temperature rise. Temperature results were expressed as the interval (days) until the temperature (oC) rise above the reference temperature, and the sum of the mean daily temperature rises above the reference temperature for the first 5 days. The results for pH were expressed as the interval in days until the pH rose by 0.2 or more above its starting value, the interval until it reached its maximum value and the maximum rise in pH.
In Experiment 3.1, the aerobic stability (12 days) of silage made from an immature maize crop was assessed in triplicate following through mixing with no additive, sodium chloride (2 or 4 g/kg silage) or sulphites (Regulator; Thomas and Fontaine Ltd., Shropshire, England; 0.2, 0.4 or 0.6 ml/kg silage, each diluted with distilled water to apply in 2 ml solution/kg silage). In Experiment 3.2, maize silage (immature crop), and in Experiments 3.3 to 3.6, unwilted grass silages, had their aerobic stability over 8 days assessed in quadruplicate following mixing with no additive or sulphites (0.4, 0.8 or 1.2 ml/kg silage, each diluted with distilled water to apply in 3 ml solution/kg silage).
Results and Discussion. In Experiment 3.1, untreated maize silage was aerobically unstable (Table 3.1). Treatment with 4 g NaCl/kg brought about a modest improvement in aerobic stability, whereas increasing rates of sulphite addition clearly improved aerobic stability, with even the lower rate of application causing a marked improvement. In Experiment 3.2, untreated maize silage was again unstable (Table 3.2), the lowest rate of sulphite addition brought about a marked improvement, and this was further but similarly improved by both of the higher rates. Sulphites improved the aerobic stability of grass silages, the effects being more evident when untreated silages were aerobically least stable and being negligible when untreated silages were aerobically stable. The mean (s.d.) DM concentration and pH of the maize silage used in Experiment 3.1 was 188 (2.6) g/kg and 3.8 (0), respectively, while the corresponding values for Experiments 3.2 through 3.6 were 206 (2.5) g/kg and 3.5 (0), 230 (5.7) g/kg and 3.7 (0), 174 (1.0) g/kg and 4.1 (0), 251 (4.6) g/kg and 3.7 (0) and 181 (2.1) and 3.4 (0).
Conclusion. Thorough mixing of sulphites with silages improved aerobic stability, the effects being greatest with the aerobically least stable silages. For a comparable aerobic stability, significantly greater rates of NaCl addition than used would be required, but such rates would be undesirable in a ruminant feedstuff. Further research is required to elucidate the effects of applying a range of rates of sulphites at ensiling on aerobic stability - this should be done using grasses varying in factors such as DM concentration.
Experiment 3.7. Ruminal digestion of silage in the presence of sulphites: dose response pattern
Introduction. Silage is inherently unstable in the presence of oxygen, allowing the respiration and growth of aerobic micro-organisms. This instability is generally initiated by yeast, with other micro-organisms, including moulds, subsequently becoming active (Woolford, 1984). A dose-dependent inhibition of such instability occurred when sulphites were mixed with silage upon first exposure to oxygen (O’Kiely, 1996). Since sulphites presumably have a partial sterilising effect on micro-organisms, it was necessary to quantify the effects of sulphites on microbial fermentation in the rumen of animals eating such sulphite-treated silage. The objective of this study was to identify if inclusion of sulphites with silage in the diet of ruminants was likely to interfere with microbial digestion in the rumen.
Materials and Methods. Phase I of the Tilley and Terry (1963) in vitro dry matter (DM) digestibility assay was used as a model of ruminal digestion of silage. Six rates of sulphites (Regulator; Thomas and Fontaine Ltd.; 0, 0.4, 0.8, 1.2, 2.0 and 4.0 ml sulphites/kg fresh silage equivalent) and 3 sources of rumen liquor (wethers offered a hay/grassmeal ration or cattle offered grass silage alone or supplemented with concentrates) were used in a 6 x 3 factorial design with quadruplicate replication. The wethers (50 kg mean liveweight) were offered hay (8.15 am) and grassmeal nuts (9.30 am) and rumen fluid was collected (9.15 am) via a rumen cannula. The cattle (Friesian steers; 550 kg mean liveweight) were offered grass silage ad libitum (10.15 am), alone or supplemented with rolled barley (3 kg per animal; 8.15 am) and rumen fluid was collected (9.15 am) via a rumen cannula. Rumen liquor was collected into thermos flasks, strained through 2 layers of cheese-cloth and incubated at 37.5oC. Into each of 72 Pyrex boiling tubes (30 mm diameter x 200 mm height) was placed 0.5 g dried (40oC for 48 h), ground (1 mm screen) grass silage (in vitro DMD 774 g/kg), 40 ml buffer solution (37.5oC; pH 6.8; Na2HPO4.12H2O, NaHCO3, NaCl, KCl, MgCl2.6H2O, CaCl2.6H2O, CO2 and water) and 10 ml rumen liquor. The sulphites were pre-mixed with aliquots of the buffer as follows: 0, 22.5, 45.0, 67.5, 112.5 and 225 m l sulphites (i.e. Regulator) per 1 l buffer. This would result in the application of 0, 0.9, 1.8, 2.7, 4.5 and 9.0 m l sulphite/0.45 g silage DM, thereby achieving the target rates per 1 kg fresh silage equivalent. The stoppered tubes were stored in the dark in a controlled environment (37.5oC) incubator shaker for 48 h. They were then centrifuged (2500 rpm for 3 min) and the residue (after decanting the supernatant and washing all precipitate in crucibles) dried overnight at 102oC. pH was recorded on each of the buffer/sulphite solutions, and on the decanted supernatant following centrifugation.
Results. Buffer solutions had pH value of 6.85. The pH and temperature of rumen liquor from sheep immediately after sampling were 6.5 and 31.2oC, silage-fed cattle 6.7 and 29.7oC and silage and concentrate fed cattle 6.4 and 31.0oC. There was a significant (P<0.001) effect of dose rate on the DM digestibility (DMD) with increasing dose rate of sulphites reducing Stage I DMD (Table 3.3). This was particularly evident at the highest rate of application, and using the rumen liquor from cattle offered silage only. Increasing dose of sulphites increased the pH of the decanted supernatant (although statistically significant it was not numerically large).
Conclusion. The lower rates of addition of sulphites did not have a negative effect on in vitro rumen digestibility of silage, but there was a negative effect at higher rates of addition.
References. O’Kiely, P. (1996). Proceedings of Eleventh International Silage Conference, University of Wales, Aberystwyth, 246-247.
Tilley, J.N.A. and Terry, R.A. (1963). Journal of the British Grassland Society, 18: 104-111.
Woolford, M.K. (1984). The Silage Fermentation. Marcel Dekker, 350 pages.
Experiment 3.8-3.9. Aerobic stability of silages made from forages treated with sulphites at ensiling
Introduction. An initial experiment demonstrated that thorough mixing of a cocktail of sulphites with silage upon exposure to oxygen at feedout enhanced aerobic stability, the magnitude of the response being greater with aerobically more unstable silages (O’Kiely, 1996). For a variety of reasons, including the opportunity to prevent aerobic losses at the feed face before removal of silage for feeding, as well as because of the greater relative ease of thoroughly mixing an additive with forage at harvesting compared to at feedout, it would be highly preferable could the aerobic stability of silage be enhanced by adding the sulphites at ensiling rather than at feedout. The following experiments were designed to test this possibility.
Materials and Methods. In Experiment 3.8, unwilted grass (17 November) and physiologically immature forage maize (12 October) were precision-chopped and ensiled in laboratory silos (6 kg forage/silo; 4 silos per treatment) alone or with the addition of a cocktail of sulphites (Regulator; Thomas and Fontaine Ltd.) applied at 0.1, 0.2 or 0.3 ml/kg fresh forage (applied at 2 ml/kg forage when diluted with distilled water). Silos were stored at 15oC for 103 and 113 days, respectively, before opening, sampling and assessment of silage aerobic stability at 20oC.
In Experiment 3.9, October harvested Lolium perenne was precision-chopped at 3 dry matter (DM) concentrations (unwilted, 24 h wilt and 48 h wilt) and manually treated in the laboratory with no additive (2 ml distilled water/kg grass) or the cocktail of sulphites at 0.2 or 0.4 ml/kg grass (diluted in distilled water and applied at 2 ml solution/kg grass). Forage from each of these treatments was ensiled immediately after the addition of additives or after being stored overnight at 15oC in unsealed polythene bags. Each of the 18 treatment combinations in the above 3 x 3 x 2 factorial arrangement of treatments were repeated in triplicate, and silos were stored at 15oC for 28 days before being weighed, sampled and silages assessed for aerobic stability.
Results and Discussion. In contrast to previous results where sulphites improved aerobic stability when mixed with silage at feedout, no benefits accrued to silage aerobic stability in Experiment 3.8 when sulphites were mixed with grass (148 (2.0) g DM/kg) or maize (177 (2.0) g DM/kg) at ensiling (Table 3.4). To test this finding further, grasses of 145 (3.5), 181 (3.5) and 296 (12.4) g DM/kg were treated with 0, 0.2 or 0.4 ml sulphites/kg and ensiled immediately or after a 24 h delay. Wilting and delayed sealing both lead to a major increase in yeast counts (and also in the numbers of lactic acid bacteria (LAB)) and sulphites did not prevent the increase due to delayed sealing (Table 3.5). Sulphites did not affect LAB numbers on herbage. After ensilage, sulphites had not reduced the counts of yeast (nor did they alter LAB number). Aerobic stability of silage was influenced by both wilting and delayed sealing, but not by the application of sulphites of ensiling (Table 3.6).
Conclusion. Whereas the application of sulphites to silage at feedout had previously been shown to improve aerobic stability, this phenomenon could not be repeated when they were added to herbage at ensiling.
Reference. O’Kiely, P. (1996). Proceedings of Eleventh International Silage Conference, University of Wales, Aberystwyth, 246-247.
Experiments 3.10-3.11. Chemical composition of silages made with the application of sulphites at ensiling
Introduction. As part of the experiments by O’Kiely, Bracken and Doyle (1997) determining the effects of mixing sulphites with forage at ensiling on the subsequent aerobic stability of silage, the effects of this selective sterilant on silage fermentation were also assessed.
Materials and Methods. In Experiment 3.10, unwilted grass (17 November) and physiologically immature forage maize (12 October) were precision-chopped and ensiled in laboratory silos (6 kg forage/silo; 4 silos per treatment) alone or with the addition of a cocktail of sulphites (Regulator; Thomas and Fontaine Ltd.) applied at 0.1, 0.2 or 0.3 ml/kg fresh forage (applied at 2 ml/kg forage when diluted with distilled water). Silos were stored at 15oC for 103 and 113 days, respectively before opening, sampling and assessment of silage aerobic stability at 20oC.
In Experiment 3.11, October harvested Lolium perenne was precision-chopped at 3 dry matter (DM) concentrations (unwilted, 24 h wilt and 48 h wilt) and manually treated in the laboratory with no additive (2 ml distilled water/kg grass) or the cocktail of sulphites at 0.2 or 0.4 ml/kg grass (diluted in distilled water and applied at 2 ml solution/kg grass). Forage from each of these treatments were ensiled immediately after the addition of additives or after being stored overnight at 15oC in unsealed polythene bags. Each of the 18 treatment combinations in the above 3 x 3 x 2 factorial arrangement of treatments was repeated in triplicate, and silos were stored at 15oC for 28 days before being weighed, sampled and assessed for aerobic stability.
Results and Discussion. The grass at ensiling in Experiment 3.10 had the following mean (sd) composition: dry matter (DM) 148 (2.0) g/kg, DM digestibility (DMD) 625 (22.4) g/kg, crude protein 179 (8.4) g/kg DM, ash 223 (13.7) g/kg DM and water soluble carbohydrates (WSC) 11 (1.5) g/L. The corresponding values for the maize were 177 (2.0), 702 (15.2), 168 (4.3), 84 (4.2) and 23 (3.0). The unwilted grass ensiled in Experiment 3.11 immediately after chopping had the following composition: DM 145 (3.5) g/kg, DMD 779 (10.4) g/kg, crude protein 315 (2.7) g/kg DM, WSC 40 (4.1) g/kg DM and buffering capacity 589 (19.8) mEq/kg DM. The corresponding values after 24 h wilting were 181 (3.5), 768 (10.1), 305 (5.4), 28 (7.1) and 589 (5.5) and after 48 h wilting were 296 (12.4), 755 (9.3), 287 (6.9), 21 (2.8) and 536 (19.3).
In Experiment 3.10 (Table 3.7), grass silage underwent an acetic acid dominant fermentation while maize underwent a lactic acid fermentation. Increasing rates of sulphite addition reduced the elevated grass silage pH by increasing lactic acid and reducing acetic acid whereas with maize silage they reduced acetic and propionic acid contents, but did not alter pH. In Experiment 3.11 (Table 3.8), wilting restricted lactic acid and ethanol production but increased acetic acid and ammonia-N concentrations while delayed sealing resulted in a reduction in lactic acid content and an increase in pH and ammonia-N, acetic acid and ethanol concentrations. Sulphites did not alter silage composition.
Conclusion: Whereas sulphites did not affect silage DM, DMD or crude protein values, their effects on fermentation varied from no effect over a wide range of conditions in Experiment 2, to the reduction of acetic acid and, if the initial values were low, the increase in acetic acid, in Experiment 1. Taken together with the data of O’Kiely, Bracken and Doyle (1997) this suggests sulphites favoured a homofermentative lactic acid fermentation.
Experiment 3.12. Aerobic stability of maize silage following addition of sulphites at ensiling or feedout
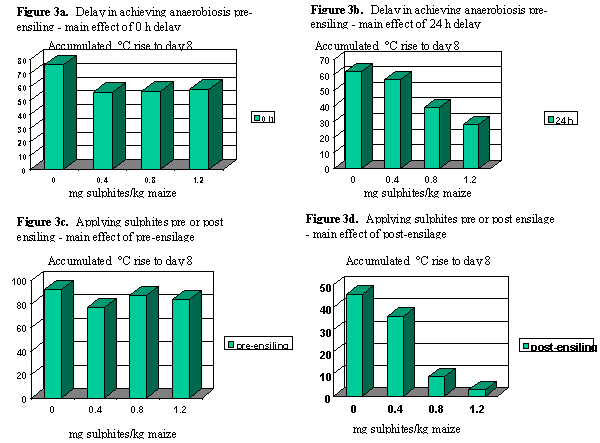
Introduction. Previous reports have shown that whereas the addition of a cocktail of sulphites to grass or physiologically immature maize silages at feedout markedly improved silage aerobic stability (O’Kiely, 1996), no benefit to silage aerobic stability was found when the sulphites were applied at ensiling (O’Kiely, 1997; O’Kiely et al., 1997). The present experiment sought to determine the effects of adding a series of rates of sulphites on the aerobic stability of maize silage of relatively high starch content when added at ensiling or at feedout, and to determine the interaction with the speed of achieving anaerobiosis at ensiling.
Materials and Methods. Approximately 500 kg precision-chop maize was obtained at harvest from a forage crop of relatively high grain content. Samples (7 kg) were placed in 64 polythene bags, with 4 bags being allocated (time 0) at random to each of 16 treatments arranged in a 2 (Level 1: addition of sulphites pre or post ensilage) x 2 (Level 2: duration of exposure to air between assigning to treatments and ensiling) x 4 (Level 3: rates of sulphite addition) factorial design: Level 1 - sulphite treatments applied at time 0 or when individual silos opened after ensilage; Level 2 - forage ensiled at time 0 or after 24 h storage (at 15oC) in unsealed polythene bags; Level 3 - sulphites added at 0, 0.4, 0.8 or 1.2 ml/kg maize. Each rate of sulphites (Regulator, Thomas and Fontaine Ltd.) addition, including non-treated, was diluted in distilled water and the solution applied at 2 ml/kg maize. Maize (6 kg) was ensiled in laboratory silos, and stored at approximately 15oC for 239 days. The aerobic stability of each silage (n=64) was assessed during exposure to air at 20oC for up to 10 days, after silo opening, weighing and sampling, and addition of sulphite treatments where appropriate.
Results. At time 0 or after 24 h non-anaerobic storage, respectively, the temperature of the forage was 14 + 1oC and 19 + 2oC. Maize silage was susceptible to aerobic deterioration at feedout (i.e. after silo opening), and the trend was for the 24 h delay in ensiling to be associated with aerobically more stable silage (Figure 3a/3b). This may have been related to the lower lactic acid and higher acetic acid concentrations evident following delayed sealing (Table 3.9). When forage maize was ensiled immediately after sulphite treatment, silage aerobic stability at feedout was not influenced by addition of sulphites. However, where achieving anaerobic conditions was delayed for 24 h, only the highest rate of sulphite addition improved aerobic stability. The biological magnitude of the latter effect was modest. Adding sulphites at silo opening gave a more marked improvement in aerobic stability. Where silos were originally sealed at 0 h, significant incremental improvements accrued to the low and medium rates of sulphite addition to silage, with similar effects being achieved with the medium and high rates. Where silos were originally filled and sealed after 24 h storage, benefits to silage aerobic stability accrued to the medium and high rates of sulphites, but not the low rate.
Conclusions: Sulphites considerably improved silage aerobic stability when intimately mixed with silage at feedout, but had a lesser (or no) effect when applied after harvesting (Figure 3c/3d). Forage maize ensiled quickly after harvesting resulted in a higher silage lactic acid:acetic acid ratio than when achieving anaerobic conditions was delayed for 24 h. This in turn was associated with poorer aerobic stability.
References. O’Kiely, P. (1996). Proceedings 11th International Silage Conference, University of Wales, Aberystwyth, p 246-247.
O’Kiely, P. (1997). Irish Journal of Agricultural and Food Research, 36: 105.
O’Kiely, P., Bracken, C. and Doyle, E.M. (1997). Irish Journal of Agricultural and Food Research, 36: 104.
Overall conclusions on sulphites to improve aerobic stability.
Addition of sulphites to silage at feedout conferred considerable aerobic stability to silages that were aerobically unstable. However, addition of sulphites to forage at ensiling conferred relatively minor benefits, if any, at subsequent feedout. The rates of addition of sulphites to silage necessary to confer aerobic stability should not negatively influence subsequent ruminal digestion.
4. A COMPUTER PROGRAM TO CALCULATE THE COST OF PROVIDING RUMINANTS WITH HOME-PRODUCED FEEDSTUFFS
Introduction: The provision of feed accounts for 85% of total direct costs on cattle farms in Ireland (Power and Roche, 1992). Purchased concentrate ingredients represent 12% of total dry matter (DM) consumption by cattle, while grazed grass and conserved grass represent 59% and 29% of DM consumption, respectively (McLoughlin, 1991). The cost of producing, storing and feeding these feedstuffs is therefore a major factor affecting profitability of cattle farms. While market price (adjusted for transport, storage and feeding costs) is the appropriate measure of cost for purchased feedstuffs, calculation of the cost of home-produced feedstuffs is often over-simplified by assigning a single universal cost to a particular feedstuff (e.g. grass silage), and by assuming that the cost of providing the feed to an animal is synonymous with its production cost. However, the true cost of providing an animal with feed is influenced by factors both outside the control (soil type, geographical location, weather conditions) and within the control (management procedures on-farm) of the farmer. To increase the accuracy of costing home-produced feedstuffs, as many of these factors as possible must be included. To facilitate this exercise, the computer program described in this paper was developed.
Program. The crops considered in the program are grass, cereals and fodder beet which collectively constitute almost all of the home-produced feedstuffs offered to ruminants in Northern Europe. Individually, their relative contributions will vary between and within countries. For each crop being studied, the physical, labour and financial inputs for producing, harvesting, processing, storing and feeding the crop, together with the resultant feed output available to be consumed by cattle, are identified and each is assigned a default value which can be accepted or altered by the user. The latter is most likely to be an advisor, specialist, consultant or researcher, but could be a farmer or farm manager.
The program is written using the C language to run on a personal computer under the MS DOS operating system. It is menu-driven and extracts default values from a crop-specific file or an overall master file. Alternatively, new default values can be added interactively, with an option on whether or not to store them permanently in the file. Movement through the program is by a series of user choices, as outlined in Figure 4.1. A context-sensitive help function is available to explain the principal data source or calculation method involved.
Crops. Grass utilisation is by grazing or by conserving as silage or hay before consumption. Conserved grass can be further categorised on the basis of sward type to include permanent swards of mixed species or reseeded swards of ryegrass and clover. Cereals are categorised by species (wheat, barley, oats or maize) to reflect their differing production procedures and, with the exception of maize, further differentiated as to whether they are winter or spring sown varieties. The small grain cereals can be harvested by conventional means or the whole crop may be ensiled, while ensilage of the whole crop is the only option considered for maize. Fodder beet is also considered and can be harvested as roots or ensiled whole.
|
Figure 4.1. Schematic outline of the program [too complicated to upload on WWW; see original document] |
Feed Outputs. The program offers the user default values for yield and quality of harvestable crop pertaining under good management practices on Irish farms. These can then be modified, as appropriate, to take account of inefficiencies at grazing and the quantitative and qualitative losses that occur during harvesting, storage, processing and feeding of non-grazed feedstuffs. The final output is the quantity of DM, digestible DM or metabolisable energy (ME) available to be consumed by the animal, per hectare of crop. Whereas the default values offered are those estimated by the authors for good management under Irish conditions, and are based on both a broad section of published literature and on practical experience, they can readily be altered by the user to suit any other set of conditions.
Table 4.1 summarises the default values offered when predicting feed output for the various crops. For forages, DM digestibility (DMD) is measured as described by Tilley and Terry (1963) and ME concentration is predicted from the equation: ME (MJ/kg DM) = ( (DMD (g/kg) - 61) x 0.011) + 3.2 (MAFF, 1984).
The values predicted for the yield and quality of grass harvested for making silage are calculated using the equations shown in Table 4.2 (derived from five years unpublished field plot data of O’Kiely), with the cutting date and regrowth interval being chosen by the user; the user may replace these predicted yield and digestibility values with values of their own choice. The remaining default values are shown in Table 4.1. Fresh weight losses during wilting are calculated, and hay as a feedstuff is assumed to have a DM content of 820 g/kg.
The main defaults for cereals grown for grain production are shown in Tables 4.1 and 4.3. Where whole crop cereal is ensiled, a ratio of 1 kg grain DM: 0.9 kg straw DM is assumed. Default values for fodder beet are presented in Table 4.1.
Farm Inputs. Input costs are classified under the headings of production and harvesting, storage, processing, feeding and ancillary costs, as shown in Figure 4.2. Standard contractor charges and material costs are used as defaults for all operations and materials, respectively (Teagasc, 1995).
Grass. For grazed grass, silage and hay, there is an option to spread the costs of reseeding over a number of years, the default value being 20 years. Similarly, the costs of lime application are spread over the default interval of 10 years (i.e. 10% of the cost attributed to any crop in one year). Fertiliser, to supply elements such as nitrogen, phosphorus and potassium, can be selected to be spread as individual elements or as mixtures of elements, and the contribution of animal slurry spread on the land can be included. Options are offered for storing conventional silage on a concrete base alone, or with the inclusion of walls and a roof as optional extras, with the default set to depreciate their cost over 20 years. Calculations for baled silage are based on storage on a serviced concrete base together with a protective netting cover, while hay is assumed to be stored in a roofed shed, with its cost depreciated over 30 years.
Cereals. Grains can be stored in systems involving low volume ventilation, propionic acid treatment, bulk drying or high temperature drying (Lenehan, 1986) or the whole crop can be stored as silage; the only option for maize is storage as silage. Default values are also included to accommodate the costs of rolling or grinding grain.
Fodder beet. Fodder beet roots are assumed to be stored on a concrete base, ventilated and protected from rain. Cleaning and slicing costs can also be included or alternatively, the option of ensiling and storing whole crop fodder beet can be costed.
The cost of bringing each feedstuff from its storage site to the feed manger is calculated within the program for each crop, using defaults based on standard labour and machinery charges (Teagasc 1995), except for grazed grass where a herding charge (time involved in moving animals from paddock to paddock) is used. Ancillary charges are included for a range of factors, including a land charge and interest. For most crops, the full annual fixed costs are charged against the crop, whereas for silage and hay, fixed costs for a cut are calculated in proportion to the duration of the entire growing season occupied in growing the crop for that cut. Where crops are eligible for European Union Area Aid, or where a by-product such as straw is produced, the program makes appropriate allowances to the returns per ha.
Applications. This computer program offers the opportunity to compare the economic value of a range of home-produced feeds, using a rapid, standardised procedure which also retains flexibility. The data in Table 4.4 show the value of well-managed grazed grass in Ireland, the important role of grass silage and the positive impact of European Union Area Aid on the competitiveness of cereals and cereal silage. The absence of EU Area Aid reduces the attractiveness of fodder beet roots, while maize is an attractive option for the minority currently capable of consistently achieving high yields. In general, home-produced feedstuffs, assuming satisfactory yields, qualities and efficiencies are achieved, can be much cheaper sources of nutrients for farm livestock than purchased compound rations (Table 4.4). Within each feedstuff, the program demonstrates a considerable range in costs, resulting in an overlap between different feeds. This illustrates the opportunity that exists to reduce significantly the costs of providing livestock with home-produced feed on individual farms. In addition, the magnitude of the range in costs for each feedstuff highlights the importance of assigning the appropriate costs to each feedstuff when formulating rations for livestock on a least-cost basis. However, because of differences between feedstuffs in the efficiency with which digestible DM and ME are utilised by cattle (AFRC, 1993), the ultimate aim is to use net energy as an index of the nutritive value of feedstuffs. The program could be adapted by the user to incorporate such an alteration whenever the necessary data were available.
Land charge. The program can be used to show the effects of including or increasing the cost of specific inputs on the unit cost of providing feed. For example, the inclusion of a land charge when determining the cost of providing home-produced feeds to animals, either as an opportunity cost incurred by not utilising the value of the land to earn an alternative income, or as the difference between producing the crop from owned or rented land, can lead to considerable increases in the unit cost for all crops (Table 4.5).
Note:
Grazed grass. Environmental factors outside the farmer’s control can lead to considerable variation in annual yield, and consequently variation in the unit cost of feeding herbivores. Variation in weather conditions from year to year in Ireland can result in grass yields deviating by up to 20% from the 20-year average (Brereton, 1995). Because the input costs of grass are essentially the same each year, the unit cost of feeding animals with grazed grass would range from about Ł40 to Ł60/t digestible DM, depending on weather conditions, with a mean of Ł48/t digestible DM. Similarly, the 20-year average annual grass yield will vary on free-draining sites due to the climatic effects associated with geographic location (Brereton, 1995). Consequently, pastures in the south-west of Ireland can be expected to produce more grass than those in the north-east (15.5 versus 10.5 t DM/ha), and have higher costs associated with higher rates of phosphorus and potassium replenishment. Thus, the unit cost of grazed grass within a defined system can vary due to geographic location from Ł35 (south-west) to Ł51 (north-east)/tonne digestible DM. Yields on wet soils in the west of Ireland may be reduced by 2 t DM/ha compared with adjacent well-drained soils, while the comparable difference between soil types in the east is about 1 t DM/ha (Brereton, 1995). The unit cost, as calculated by this program, of providing grazed grass within a common system in a given year, increases due to wet soils from Ł45 to Ł53/t digestible DM in the west of Ireland and from Ł45 to Ł49/t digestible DM in the east.
The two critical components of grazing management are the production and efficient utilisation of sufficient quality grass throughout the year. In studying the marginal financial response to sequential increments in fertiliser nitrogen (N) application, the program indicates that, even though grazed grass is a relatively cheap feed source, alternative feed sources should be considered, on economic grounds, in preference to increasing N application rates above 250 kg N/ha, because of the declining grass yield response at higher rates of N application. Even under good grazing management, ruminants will not consume all of the grass produced. With excellent grazing management, beef cattle are likely to consume about 74% of annual grass DM produced, while under poor grazing management the value is likely to be below 64% (O’Riordan, 1996). Cattle are quite selective in their grazing behaviour, tending to choose the highest quality grass available. Where grassland management is excellent cattle may consume grass of 780 g digestible DM/kg DM, whereas with bad grazing management it may be 720 g digestible DM/kg DM. An analysis of these two extremes of grazing management shows that the unit cost of grazed grass can range from Ł48/t digestible DM (74% grazing efficiency and 780 g digestible DM/kg DM) to Ł60 t/ digestible DM (64% grazing efficiency and 720 g digestible DM/kg DM), demonstrating the financial reward for achieving high standards of management.
Grass silage. The program has been used to examine, in Table 4.6, the effects of variation in yield and digestibility of grass for ensiling on the unit cost of silage. These results indicate that within a given system, light yielding crops of conventionally-harvested grass produce a very expensive feedstuff, and that the normal range in yields will have a more dramatic effect on feed costs than the normal range in digestibility.
As the harvesting date of grass for ensilage is delayed, yields increase while digestibility declines, particularly with the primary growth (Table 4.2). The balance between quantity and quality is such that the unit cost of producing digestible DM or ME also declines. However, the optimal harvesting date has to be calculated for individual farms, to take account of factors such as the reduction in voluntary intake of silage associated with a decrease in digestibility, the stocking rate on the farm, the cost and availability of alternative feedstuffs, the desired level of individual animal performance and the sale value of animal produce. Consequently, on many spring-calving dairy farms, the optimal balance between yield and digestibility will be achieved with successive silage harvests on dates such as May 28 and July 18 rather than either a more intensive three-cut regime (e.g. May 20, July 1 and August 12) or a more extensive two-cut regime (e.g. June 10 and August 12).
The model can be used to quantify the effects of a wide range of other factors on the cost of providing silage for animals. For example, it shows that when dealing with high yields of unwilted grass, single and double chop harvester systems can, in general, produce digestible DM or ME more cheaply than precision-chop systems, while precision-chop systems in turn can produce silage more cheaply than the wrapped big bale system. However, as yields decrease and if effective wilting is carried out, the big bale system becomes progressively more competitive. In practice, factors other than the unit cost of providing digestible DM or ME will often determine which system is used.
Silos represent an expensive capital asset, with a concrete base being much cheaper to construct than a walled, roofed silo. When depreciated over 20 years however, the unit cost of feeding animals with a standard silage stored on either a concrete base, or in a walled or a walled, roofed silo was calculated to be Ł107, Ł116 and Ł125/t DDM, respectively, corresponding to Ł7.3, Ł7.9 and Ł8.5/GJ ME. On the other hand, reducing conservation losses (field and silo combined) from 300 to 200 g/kg DM reduces the unit cost of providing DDM from Ł122 to Ł107/t, equivalent to reducing the cost of providing ME from Ł8.3 to Ł7.3/GJ. In each of the above examples, a different set of absolute values would be obtained were the input and output assumptions altered, but the relative effects of the harvesting regimes, silo types and conservation efficiencies being contrasted would remain intact.
Cereals. Cereal grain has traditionally been considered an expensive feed source compared to forage. Table 4.7 shows the cost of providing DDM or ME to ruminants for each of the major cereal grain types, both with and without EU Area Aid support. The yields used are 5-year averages for grain of 200 g moisture/kg, grown on suitable soils with good management (A. Doyle, personal communication, 1994). The unit costs are lower for the higher yielding cereals, and EU Area Aid has a major impact on making the unit cost of grain more attractive. European Union Area Aid also reduces the relative difference in unit cost between cereal types.
Fodder beet. On a full cost basis (excluding land charge), the cost of producing DDM or ME in the form of fodder beet roots is quite attractive, but due to potentially high storage and feeding costs, it can become a very expensive feedstuff for cattle (Table 4.8). Obviously, the particular circumstances prevailing on individual farms can alter the situation considerably. Furthermore, the high intake characteristics associated with fodder beet can mean excellent levels of animal performance.
Conclusions. As far as the authors of this report are aware, this program is unique in that it measures the full cost of delivering feed to the animal as required. In developing the system, information was gathered from a wide variety of sources, and the collation of these data into an easily accessible database has, in itself, been useful. The system is simple to operate and requires minimal computing facilities. It can quickly cost feeds under chosen conditions and can analyse the effects of alternative management decisions relating to operational or material inputs or the effects of varying quantitative or qualitative aspects of output.
In this report it has been used to assess the relative costs of a number of home-produced animal feeds under certain chosen conditions, and a range of issues have been assessed using sensitivity analysis. This model could also be used as a research tool in designing various prototype systems for farms, or to identify the priority research areas from which technological improvements could lead to the greatest economic benefit. Alternatively, it can be used as an aid to policy strategies in advisory or consultancy work. In individual farm situations, quantification of feed outputs would be necessary, and the real, specific costs of both variable and fixed inputs pertaining would be calculated and used. These could substantially alter the relative economic attractiveness of some feedstuffs compared to the generalised situations described in this report.
References. Agriculture and Food Research Council (1993) Energy and Protein Requirements of Ruminants. An Advisory Manual prepared by the AFRC Technical Committee on Responses to Nutrients. CAB International, Wallingford, UK, 159 pp.
Brereton, A.J. (1995) Regional and year to year variation in production. In: D.W. Jeffrey, M.B. Jones and J.H. McAdam (Editors), Irish Grasslands - Their Biology and Management, Royal Irish Academy, Dublin, pp 12-22.
Lenehan, J.J. (1986) Grain drying and storage-principles of drying and storing combinable crops. An Foras Taluntais, Dublin, 59 pp.
McLoughlin, M. (1991) Feed balance sheets for Ireland 1983/84 to 1988/89 - supply, demand and allocation. Paper presented to the Agricultural Economics Society of Ireland, Dublin, December, 73 pp.
Ministry of Agriculture, Fisheries and Food (1984) Energy allowances and feeding systems for ruminants. Reference Book 433, Her Majesty’s Stationery Office, London, 85 pp.
O’Riordan, E.G. (1996) Beef production from grazed grass and grass/clover swards. Proceedings of Agricultural Research Forum, University College Dublin, pp 117-118.
Power, R. and Roche, M. (1992) Cattle in the national farm survey. Teagasc, Dublin, 42 pp.
Teagasc (1995) Management Data for Farm Planning (1995). Teagasc, Dublin, 200 pp.
Tilley, J.M.A. and Terry, R.A. (1963). A two-stage technique for the in vitro digestion of forage crops. Journal of the British Grassland Society, 18: 101-111.
1. Silages made from the annual output of an old permanent grassland sward of high agronomic quality, and conserved as unwilted silage, were compared in terms of beef carcass output per hectare with comparable herbage from perennial or Italian ryegrass reseeds. Beef carcass output per hectare was in the order Italian ryegrasss > perennial ryegrass > old permanent grassland. The carcass output differences arose from the cumulative effects of changes in components such as yield, digestibility, ensilability, conservation efficiency, silage quality, silage intake, feed conversion efficiency and carcass gain.
2. The economic benefits (margin over feed) were in the order perennial ryegrass > Italian ryegrass > old permanent grassland. The cost of the additional harvest of Italian ryegrass, the apparently higher in-silo losses and the need for more frequent reseeding, militated against the Italian ryegrass.
3. Conclusions were also drawn on optimum harvesting intervals, fertiliser nitrogen application dates and rates, and the justification for reseeding under high input and low input farming systems. Reseeding silage swards is more readily justifiable under intensive farming conditions, but its sustainability still depends on the quality and productivity of the old pasture being replaced, and on the new ryegrass reseed being managed in a manner that would maintain its long-term dominance in the sward and efficiently convert its output to animal product.
4. Difficult silage-making conditions reflected in grass crops of low water soluble carbohydrate (and dry matter) concentration and/or high buffering capacity still occur frequently in Ireland. A series of experiments using laboratory silos showed no beneficial effect of homofermentative lactic acid bacterial (HmLAB) inoculants on preservation or conservation variables under such conditions. Similarly, two experiments using farm-scale silos showed no production response by beef cattle to the use of a similar (HmLAB) inoculant under difficult ensiling conditions. In contrast, benefits did accrue from the use of formic acid. The use of strains or species of HmLAB selected from Irish silages, or the inclusion of selective sterilant with the HmLAB inoculant, did not improve the response. Overall, the data indicate that HmLAB inoculants should not currently be recommended as silage additives under difficult ensiling conditions.
5. A series of experiments studied the ability added sulphites to improve the aerobic stability of silage. They conferred considerable stability when mixed with aerobically unstable silage at feedout. However, their addition at ensiling conferred relatively minor or no benefits at subsequent feedout.
6. Cost of feeding cattle with home-produced feeds. A computer program was developed to facilitate the rapid and standardised calculation of the cost of providing home-produced feedstuffs for ruminants in Ireland. The user, most likely an advisor, researcher, specialist or consultant, is guided via a menu system through a sequence of options (for each of which defaults are provided that can easily be replaced) to choose the management, physical, biological and financial components of producing, harvesting, storing, processing and feeding various crops, as appropriate. Direct and indirect (e.g. land charge) costs, inherent efficiencies, subsidies (e.g. European Union area aid), etc. can all be taken into account. Feeds are costs per unit of dry matter (DM), digestible DM or metabolisable energy (ME) available for consumption. Among the unit costs determined were those of providing ruminants with grazed grass, individual harvests of grass silage or hay, maize silage, fodder beet roots (or whole crop silage) and wheat, barley or oaten grain (or whole crop silage).
|
Top | Acknowledgements | Publications | Tables | Teagasc [Irish Agriculture & Food Development Authority] Homepage |